Overview: Bipedalism is a defining characteristic of modern humans that evolved over millions of years. Therefore, identifying evidence for bipedalism in the fossil record can help determine what selective pressures may have affected human evolution. This lab examines the significance of bipedalism, anatomical adaptations exhibited by hominins, and discusses possible climatic influences on bipedal evolution. Look over the table and questions at the end of this page, and then read through the lab information, completing the table as you go. (This lab has been adapted from eFossils.org)
Lab Objectives:
- To understand the significance of identifying bipedal evidence within the fossil record.
- To learn the morphological adaptations associated with bipedalism.
- To become familiar with the environmental and behavioral pressures that may have affected bipedal locomotion.
What is Bipedalism?
Bipedalism refers to locomoting (e.g., walking, jogging, running, etc.) on 2 legs. It is not uncommon to see animals standing or walking on 2 legs, but only a few animals practice bipedalism as their usual means of locomotion. Animals, including chimpanzees and gorillas, that assume bipedalism on a temporary basis in order to perform a particular function practice a form of locomotion called facultative bipedalism. For example, octopi sometimes walk bipedally in order to camouflage themselves from predators1 . The octopus piles 6 of its 8 limbs on top of its head, assuming the shape of a drifting plant, and then uses the 2 remaining limbs to quite literally walk away. As for quadrupeds (animals that move on four limbs), it is not uncommon to see antelope standing on their 2 hind limbs while supporting themselves on their forelimbs when reaching for food in high branches. As we’ve watched in class, chimpanzees and gorillas have been documented walking on 2 legs in order to carry things with their hands.
Watch the two short clips on the right to see examples of facultative bipeds in action.
Habitual bipedalism, which is rare, is the form of bipedalism that is assumed as a regular (i.e., habitual) means of locomotion. Today, very few mammals (e.g., humans and kangaroos) demonstrate habitual bipedalism. However, many early hominins (modern humans and all their bipedal ancestors) show a combination of primitive and novel adaptations that suggest these species utilized bipedalism but still engaged in arboreal behaviors. While habitual bipeds most frequently engage in bipedal locomotion, they were also adept at navigating arboreal environments when they needed to. Humans engage in a more restricted form of habitual bipedality, known as obligate bipeds. In this locomotion type, we are restricted to be able to ONLY move bipedally; we have lost our ability to move around successfully in trees. Humans are the only living primate to engage in habitual or obligate bipedalism.
Bipedalism Geological Age and Climate
Around 7 or 8 million years ago (mya), the earth’s climate underwent a dramatic cooling event which lowered land and ocean temperatures. Growth in the Antarctic ice cap during this time resulted in a dramatic drop in sea levels, including the Mediterranean Sea. As a result of these sea level changes in the Mediterranean, water source availability within nearby continents like Africa were severely limited. Thus, the extensive, moisture-dependent forests of these continents were reduced as their water sources dried up. This shift toward less dense forests and the subsequent growth in woodland environments may have been a driving force for bipedal evolution in hominins.

Recent studies have determined that the early ancestors of humans probably lived in some sort of wooded habitat, perhaps a woodland savanna. Climbing trees in search of food or to escape predators would have been a common behavior for organisms living a wooded or forest environment, and it is possible that early bipedal ancestors retained features (i.e., long arms, and curved fingers and toes) that were adapted to arboreal locomotion. In fact, some of the early hominin fossils do exhibit morphological adaptations conducive to tree climbing.
Artistic renderings of possible wooded savannah environments of early hominins


If bipedalism is one of the defining characteristics for hominins, then bipedal characteristics may be used to pinpoint the first appearance of hominins. To put it another way, although the DNA evidence suggests that apes and humans shared a common ancestor sometime between 7 and 8 mya, characteristics of this shared ancestor remain somewhat debated. The identification of early bipedal adaptations within the fossil record may help to identify this shared ancestor, or perhaps help to determine what characters would be expected in this ancestor. Therefore, understanding the evolution of bipedalism remains an important study in the story of human origins.
Morphology of Bipedalism
Bipeds have adapted a number of interdependent morphological characteristics that solve challenges posed by habitual bipedalism. These anatomical adaptations evolved over millions of years and differences exist between earlier and later hominin species (i.e., Australopithecus, Paranthropus, and Homo). Australopith and paranthropine evolution represents a notable step in the evolution of humans because these species are among the earliest hominins known to have evolved the adaptation of bipedalism.
All of the anatomical adaptations necessary for habitual bipedalism can be found in the fossil record. By reconciling the fossil evidence with the geologic time scale, it is possible to hypothesize about the evolutionary origins of bipedalism. The following is a detailed discussion of each morphological adaptation for habitual bipedalism.
Cranium:

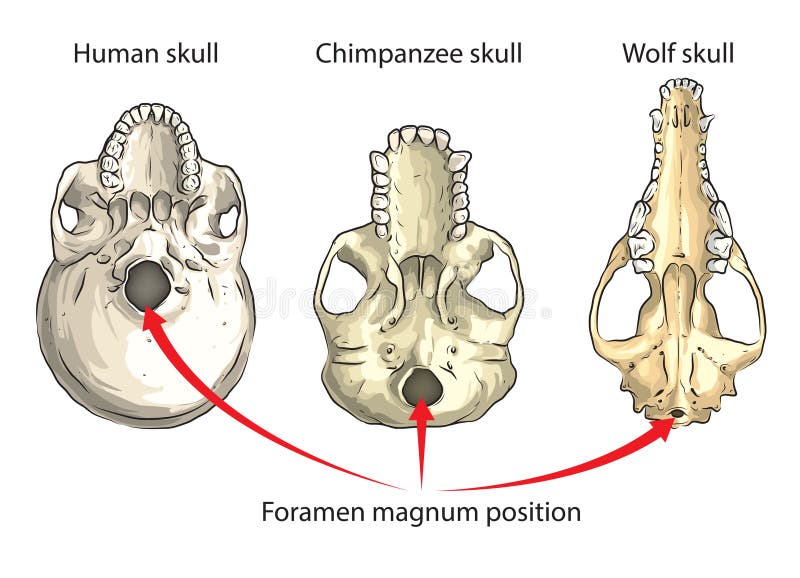
The placement of the foramen magnum, the large hole on the cranium through which the spinal column passes, is directly related to the orientation of the cranium. Consider that a primate holds their mandible (or chin) parallel to the ground. In a quadruped, the spinal column also runs parallel to the ground so the foramen magnum is more dorsally placed (i.e., toward the back of the cranium). In a biped, the spinal column runs perpendicular to the mandible and the ground. The foramen magnum is located more inferiorly (more on the bottom of the cranium). Australopiths have a more inferiorly placed foramen magnum.
Lumbar vertebra:
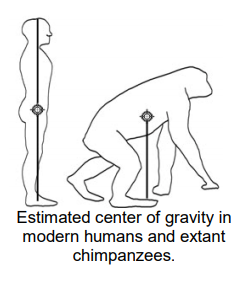
Maintaining balance is critical when walking on two legs. In part of the walking cycle, a biped must balance on one leg while lifting the other foot off the ground and swinging it forward. In most quadrupedal hominins, the center of gravity is located near center on the torso. In modern humans, the center of gravity is closer to the center of the pelvis. As the legs alternate swinging forward during the walking cycle, the center of gravity shifts from one side of the pelvis to the other, making a pattern similar to the figure 8. The lumbar curvature on the spine helps to bring the center of gravity closer to the body’s midline and above the feet.
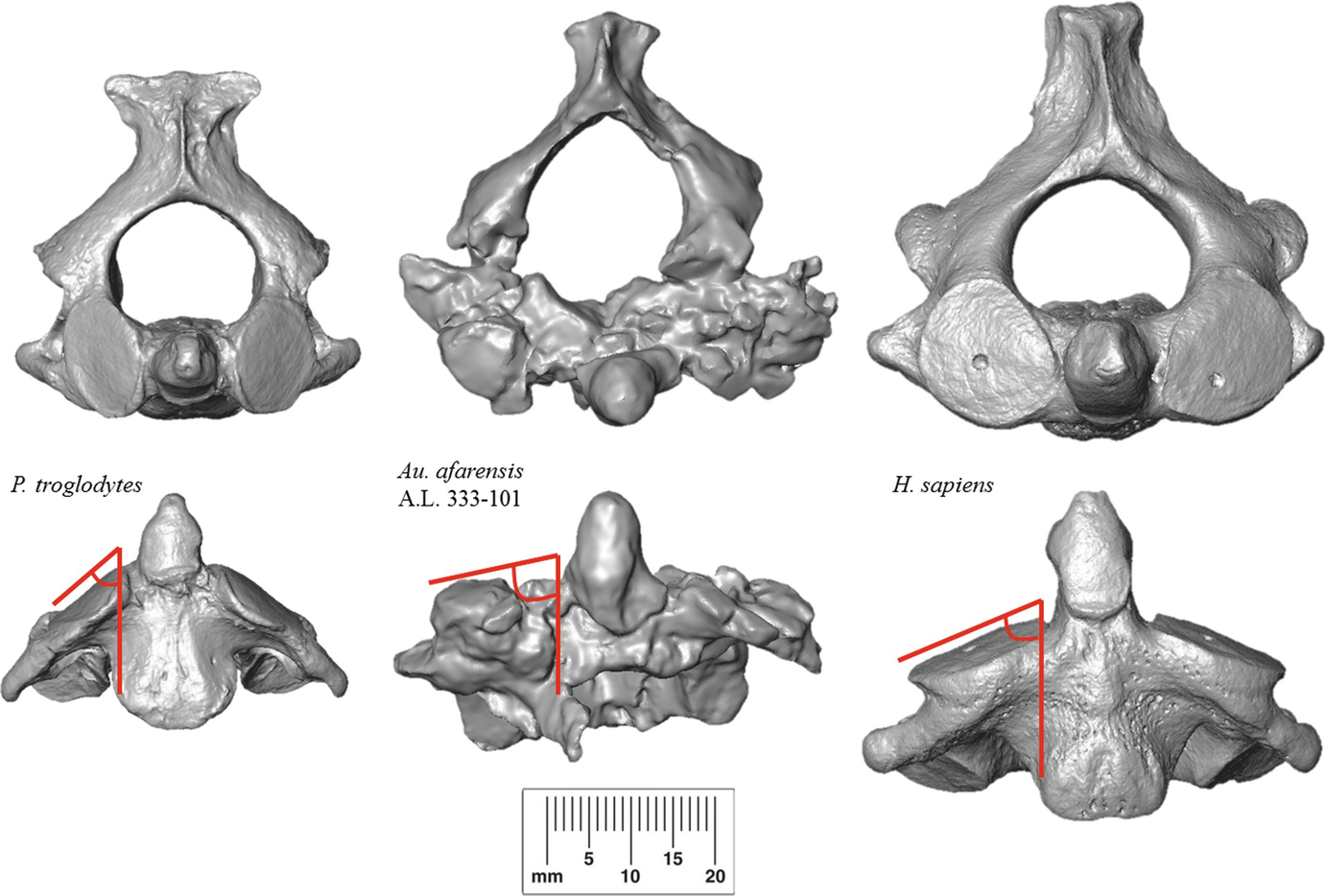
The number and size of the lumbar vertebrae in humans is different than in apes. Humans usually have 5 comparatively larger lumbar vertebrae. Most large apes typically have 4 lumbar vertebrae that are relatively smaller than human lumbar vertebrae. The greater number and size of the vertebrae forms a more flexible lower back that permits the hips and trunk to swivel forward when walking. Because the ape lower back is less flexible, the hips must shift a greater distance forward with each step when an ape walks bipedally.
Australopithecus lumbar vertebral bodies were broad for effective weight transmission from the upper body to the pelvis. Australopiths had 5 or 6 lumbar vertebrae that articulated to form a distinctive lumbar curvature, similar to the morphology of modern humans.
TASK: Zoom in on the vertebrae of the gorilla, chimp, and human below to look at the similarities and differences, and look at the image comparing the Pan troglodytes (chimp), Australopithecus afarnesis, and Homo sapiens sacral vertebrae.
The Pelvis:
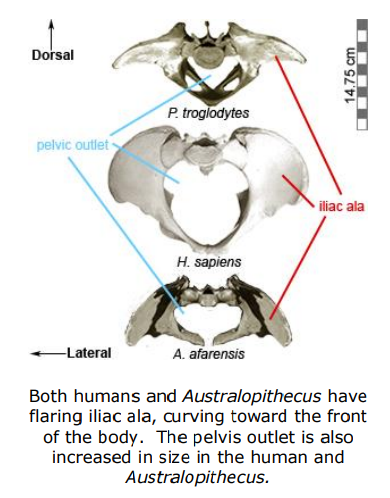
Since bipedalism requires special adaptations, the orientation (and thus the function) of the gluteal muscles, is different in bipedal humans and quadrupedal apes. In apes, the flat portion of the iliac ala is roughly parallel with the plane of the back, while in humans the iliac ala is shifted laterally and flares more on the sides. This relatively lateral orientation of the alae in humans abducts (i.e., move away from the body) the hip joint. In turn, the gluteal muscles act to stabilize the area by preventing the hip on the supported side (the standing leg) from collapsing toward the unsupported side (the swinging leg). In apes, these muscles are attached relatively dorsal (i.e., more toward the back and less on the sides) and act as hip extensors, which move the leg backward when the primate takes a step .
The australopith pelvis exhibits widely flaring iliac ala. This flare is a critical component of the lever system of the hip and acts to increase the mechanical advantage of the lesser gluteals by increasing their lever arm. However, the lateral flare of the australopith ala is more pronounced than typically seen in modern humans. Shape similarities between the australopith and modern human pelves indicate that Australopithecus was fully bipedal. However, the unique morphology seen in australopiths suggests the species did not utilize the modern gait seen in later Homo.
The modern human pelvis has relatively larger hip joints and larger pelvic outlet relative to australopiths or modern apes. These differences appear to be a compromise between two functional needs: 1) efficient bipedalism; and 2) allowing enough space for wide shouldered, large brained infants to pass through the birth canal.
In bipeds, the hips support and balance the weight of the torso during locomotion. However, as the size of the pelvic outlet increased, the hip joints were repositioned relatively further from the center line of the body. As a result, more force is exerted on the hip joint as the joint (acetabulum and femoral head) moves further away from the body’s center of gravity, and thus affects stability as the weight of the torso presses downward toward the middle of the body. This issue is resolved through several adaptations in the pelvis and femur. In the pelvis, an enlarged hip joint allows more stress to be absorbed and accommodates a larger femoral head.
Femur:
The entire weight of the torso is transferred through the legs and into the feet during bipedal standing and walking. Therefore, the femur in bipeds is one of the most critical links between the pelvis, vertebral column, and lower legs. The femur is also the distal attachment point for the gluteal muscles that provide the propulsive force for locomotion.
The rounded femoral head articulates with the pelvis at the acetabulum (hip joint). The femoral shaft is generally straight, ending in two bulbous condyles. These condyles are larger and more elliptical in bipeds when compared to the relatively smaller and rounder condyles seen in quadrupeds. The distal end of the femur articulates with the tibia (lower leg) and patella (knee cap) at the knee joint.
The amount of force exerted on the hip joint and the femoral head increases as the acetabulum moves further away from the body’s center of gravity. The size of the femoral head reflects the amount of force absorbed at the hip joint. A femoral head with a larger diameter is able to absorb more stress. Another adaptation to counteract the increased stress on the hip joint is a longer femoral neck, which increases the mechanical advantage of the lesser gluteal muscles by lengthening their lever arm.
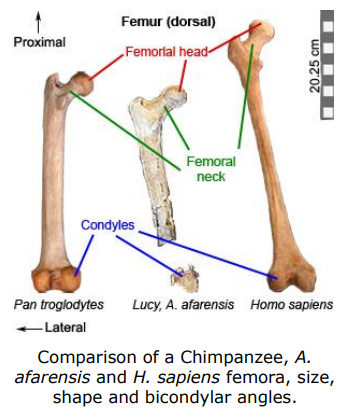
Australopithecus has a relatively smaller femoral head and longer femoral neck compared to later Homo who have a relatively and absolutely larger femoral head.
Knee (Distal Femur and Proximal Tibia):
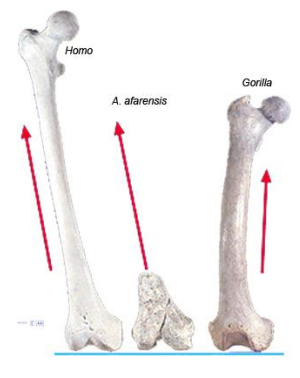
A critical adaptation for efficient bipedalism relates to the need to keep the body’s center of gravity balanced over the stance leg during the stride cycle. Birds solved this issue by having the entire leg (from the hips all the way to the feet) as close as possible to the body’s center line. In humans, whose hips are wide apart, the shaft of the femur is angled so that the knee is closer to the body’s midline than the hips. This angle is called the bicondylar angle, and the resulting knee joint is referred to as a valgus knee. The effect is to bring the knees closer together, placing the feet directly below the center of gravity.
Compared to modern humans, an ape femur is almost vertical within a horizontal plane. In quadrupeds the positioning of the center of gravity during locomotion is less critical since the quadruped is usually supported by 2 or more legs during the stride cycle rather than just 1 as with humans. Australopiths have a human-like bicondylar angle.
Fingers:

Among hominins, the degree of curvature observed in the phalangeal shaft correlates with the frequency of arboreal behavior. Species that spend a lot of time grasping or suspending from curved branches have dramatically curved fingers and toes which allows for a more powerful grip. Non-arboreal primates, such as humans, have relatively flat manual and pedal phalanges, an adaptation reflecting a lack of regular arboreal activity. This, in turn, has facilitated the evolution of precise hand movements necessary for making and using tools. Highly curved phalanges reduce the capacity for precision grips.
Australopith phalanges are intermediately curved between those of modern humans and great apes, suggesting that climbing and arboreal behavior continued to play some role in the lifestyle of these early hominins.
Arms and Legs:
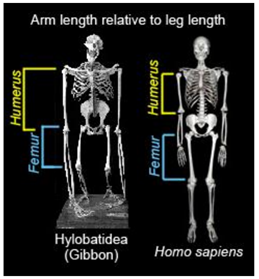
Most quadrupedal and arboreal primates have either longer arms relative to their legs, or arms and legs of equal length. Most bipeds have relatively longer legs than arms. Based on this information, it is possible to estimate the positional behavior of a species by calculating the humerofemoral index. This index is the length of the humerus divided by the length of the femur, multiplied by 100: (humerus length ÷ femur length)*100.
Results of the humerofemoral index calculate the overall body proportion of an organism which can then be compared to others. The higher the index value, the longer the arms and the more likely a primate is to be arboreal. Most arboreal primates have ratios close to 100. For example, the mean ration for the common chimpanzee is 97.8. Humans average a lower ratio at approximately 71.8. The ratio of the famous A. afarensis “Lucy” is intermediate between modern humans and chimpanzees at 84.
Feet and Toes:
Humans have the most distinctive feet of all the apes. Since only the hindlimbs (or lower limbs) are used for propulsion, the body’s entire torso weight (all of the forces generated by running, walking, and jumping) pass through only 1 foot at a time as the biped moves between the swing and stance phases of locomotion. As a consequence, the foot anatomy must be robust enough to accommodate these forces, while also providing efficient toe-initiated push-off for propulsion. As an example, the hallux (i.e., big toe) in humans is much larger and more robust than the other four toes.
The calcaneus, or heel bone, is also relatively large and robust in humans compared to chimpanzees, especially the posterior portion known as the calcaneal tuberosity. As the first foot bone to contact the ground during the stride cycle, the robust size of the calcaneus provides stability and helps to absorb the high forces encountered during heel strike. In addition, the shape of the calcaneus provides attachment points for strong ligaments that run from the arch of the foot to the tibia. These ligaments add support, creating a double arch system that helps to absorb stress as the foot hits the ground.
Part 2: Fossil Evidence of Bipedalism
The fossil record offers clues as to the origins of bipedalism, which in turn helps us to identify those species ancestral to modern humans. One of the most abundant sources for early bipedalism is found in Australopithecus afarensis, a species that lived between approximately 4 and 2.8 mya. Au. afarensis postcrania fossils clearly shows hip, knee, and foot morphology distinctive to bipedalism.
In addition to the postcranial material, Au. afarensis also left behind a 27 meter long set of footprints known as the Laetoli Tracks in Tanzania. Approximately 3.7 mya, 3 Au. afarensis individuals walked through a muddy layer of volcanic ash that preserved their foot prints after the ash hardened. From the Laetoli tracks it is clear that Au. afarensis walked with an upright posture, with a strong heel strike and follow-through to the ball of the foot, with the hallux making last contact with the ground before push-off. Interestingly, the prints provide evidence of a slight gap between the hallux and the other toes. This gap suggests that even though the hallux was not fully divergent, it was also not yet fully adducted as seen in modern humans.
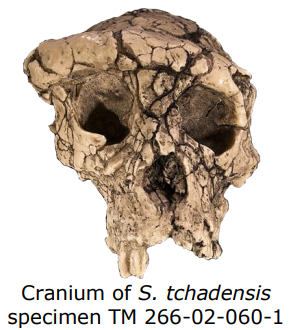
Though australopiths material offers a strong case for habitual bipedalism, earlier hominins dating as far back as 7 mya also provide exciting evidence for early bipedalism. The oldest known hominin to show definitive bipedal adaptations is the extinct species Orrorin tugenensis that dates to 6 mya. A femur and tibia recovered in Kenya and assigned to O. tugenensis exhibits feature typical of bipeds, including a bicondylar angle. However, a 7 mya fossil discovered in Chad in 2001, known as Sahelanthropus tchadensis, exhibits a more inferiorly positioned foramen magnum consistent with bipedalism, rather than the relatively dorsal position seen in quadrupeds. Both S. tchadensis and O. tugenensis lived on lake margins near the edge of woodlands and grasslands. No postcranial material has been associated with S. tchadensis, but if proven to be bipedal, this may substantiate the hypothesis that bipedal evolution was influenced by climate trends beginning in the late Miocene (i.e., a geologic epoch that dates between 23 and 5.3 Mya).
About 2 million years younger than O. tugenensis is a hominin known as Ardipithecus ramidus that dates to approximately 4.4 Ma. Known as Ardi, Ar ramidus material exhibits a mosaic of primitive and derived features, including a fully abductable hallux (primitive), relatively inflexible midfoot (derived), arms and legs of similar proportions (primitive), relatively broad iliac ala (derived), and an inferiorly placed foramen magnum.
The oldest evidence for australopith bipedalism is found in the species Australopithecus anamensis (4.2 to 3.9 mya). Found in Kenya, Au. anamensis most likely lived in a wooded savanna. Fossil evidence for this species includes a preserved tibia that exhibits bipedal characteristics such as a right angle between the shaft and the proximal surface, and proximal articular condyles of nearly equal size. An abundance of the younger species Au. afarensis (4 to 2.8 mya) and Australopithecus africanus (3 to 2 mya) fossils also show clear signs of bipedalism, including a bicondylar angle, an anteriorly placed foramen magnum, laterally flaring iliac blades, longer femoral necks and heads, and the presence of a lumbar curve. Though Au. afarensis seems to have originated in Ethiopia and Au. africanus is found only in South Africa, both of these species lived in open habitats, possibly wooded savanna areas near a lake.
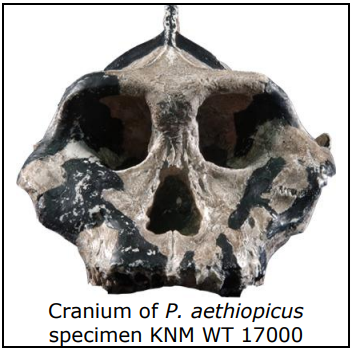
Paranthropines are larger and more robust than australopiths, but have similar postcranial morphology, including bipedal adaptations similar to Australopithecus. The oldest paranthropine was found in Ethiopia and is known as Paranthropus aethiopicus (2.6 to 2.5 mya). Although postcranial material is scarce, a possible P. aethiopicus calcaneus may exhibit bipedal adaptations. The younger paranthropine species, P. robustus (1.75 to 1.5 mya) and P. boisei (2.5 to 1 mya), exhibit the same bipedal adaptations as Au. africanus, which include an inferiorly oriented foramen magnum, modern humanlike calcaneus, relatively long femoral neck, and a bicondylar angle. In addition, the hand anatomy of P. robustus implies a grip capable of tool use, while the radius of both P. robustus and P. boisei implies Paranthropus retained the ability to effectively climb trees. Paleoecological studies suggest these species were living in open woodland or savanna habitats.
All species included in the genus Homo are obligatory bipeds and show evidence of tool use, beginning with the species Homo habilis (i.e. “Handy Man”) that dates between approximately 2.6 to 1.6 mya, and continuing to the modern species Homo sapiens that dates between approximately 190,000 years ago (kya) to present .
Bipedalism vs. Brain size?
Early researchers hypothesized that brain enlargement was the first hallmark of the hominin lineage. Beginning in the mid 1800’s until the early 1900’s, almost all known fossil hominins had relatively large brains. The large brain hypothesis was falsified after the discovery of early hominin fossils exhibiting ape-sized brains and bipedally-adapted morphology.
In 1924, Raymond Dart identified the first australopith fossil, known as the Taung Child, from South Africa. This specimen belonged to the species Au. africanus and had a relatively small brain similar to the size of a modern chimpanzees. The inferior placement of the foramen magnum, Dart argued, suggested that the Taung Child was bipedal. Dart’s hypothesis that bipedalism evolved before larger brains ran counter to the scientific consensus at the time. Because of his small sample size and fragmentary remains, debate about the timing of bipedalism and brain size continued for the next 50 years.
Everything changed in 1974 when Donald Johanson found the nearly complete fossilized skeleton of Lucy, a member of the species Au. afarensis dating to 3.2 mya. Lucy was unique at that time because she was one of the first fossils to exhibit both small relative brain size and the highly derived features characteristic of bipedalism.
Another major bipedality-related discovery came in 1992, when Tim White and his team discovered the first Ardipithecus specimens in the Middle Awash area of Ethiopia. White and his colleagues gave their discovery the name Ardipithecus ramidus (‘ramid’ means ‘root’ in the Afar language of Ethiopia and refers to the closeness of this new species to the roots of humanity, while ‘Ardi’ means ‘ground’ or ‘floor’). At the time of this discovery, the genus Australopithecus was scientifically well established, so White devised the genus name Ardipithecus to distinguish this new genus from Australopithecus. In 2009, scientists formally announced and published the findings of a partial skeleton (ARA-VP-6/500), nicknamed “Ardi”, first found in 1994. The foot bones in this skeleton indicate a divergent large toe combined with a rigid foot – it’s still unclear what this means concerning bipedal behavior. The pelvis, reconstructed from a crushed specimen, is said to show adaptations that combine tree-climbing and bipedal activity. The discoverers argue that the ‘Ardi’ skeleton reflects a human-African ape common ancestor that was not chimpanzee-like. A good sample of canine teeth of this species indicates very little difference in size between males and females in this species. Ardi’s fossils were found alongside faunal remains indicating she lived in a wooded environment. This contradicts the open savanna theory for the origin of bipedalism, which states that humans learned to walk upright as climates became drier and environments became more open and grassy.
As other contemporaneous and older fossils (perhaps as old at 7 mya) are found, scientists continue to revise the bipedalism timeline. Today, the evidence undoubtedly demonstrates that bipedalism was one of the first hallmarks of the hominin lineage and may have led to many more advances. For example, one advantage of bipedalism is that the hands are freed, which allowed for the production of more technologically advanced stone tools. In turn, the production of more complex tools may have led to a higher protein diet that affected brain size.
The functional demands of bipedalism have exerted a strong influence on the postcranial skeletal adaptations of modern humans as well as extinct hominins. For example, australopiths share with modern humans many of the essential features of bipedalism such as reorganized pelvic and lower back anatomy, a valgus knee, and a relatively robust calcaneus. However, australopiths have many unique features that differ from modern humans in significant ways. Humans do not share the long ala of the ilia, the relatively smaller femoral heads, or the curved fingers and toes seen in Australopithecus. This combination of primitive and derived features leads many researchers to support the idea that australopiths engaged in a form of locomotion that was not identical to that of modern humans, including a greater amount of time engaged in climbing and suspensory behaviors. Australopithecus may, then, represent a mosaic of evolutionary adaptations for life on the ground and in the trees.

YOUR TASK
Based on the reading, identify the species (not including the genus Homo) on the table below that show evidence of bipedalism, the geological dates associated with each species, and the morphological features that demonstrate possible bipedalism. Identify any environmental or behavioral changes that may have affected adaptations for bipedal locomotion. Remember to write down only information for which there is evidence. Then answer the reflection questions.
(Supplemental links from table above)
Sahelanthropus tchadensis skull
Ardipithecus ramidus skull
Australopithecus anamensis extra information
Australopithecus afarensis Walking with Lucy
Australopithecus africanus extra information
Based on your reading and the above exercise, complete the following questions.
- What is bipedalism?
- What is habitual or obligate bipedalism? How is it different from facultative bipedalism? For what reason(s) would a quadruped use the bipedal form of locomotion?
- What are the earliest fossil hominins to show physical evidence of bipedalism?
- What are some of the theories put forward by scientist for the evolution of bipedalism? Which one do you think is the most plausible and why? Do you think there is more than one?
- What are the anatomical features indicative of bipedalism? How do those features differ from a quadruped?
- Based on the information you have recorded, did bipedalism evolve before or after an increase in brain size? In what way could brain size have been affected by bipedalism?
- What features, if any, are unique to australopiths in comparison to modern humans and chimpanzees? What might these features indicate?